Tutti gli hotel a Roma
Museo Civico di Zoologia a Roma

| MUSEO | SCIENZA | EDUCAZIONE | EVENTI | NOTIZIE | CONTATTI |


Hotel vicino Portuense

Hotel vicino Prenestino Labicano





Hotel vicino Repubblica









Hotel vicino Rione Monti




!SEOSLIDER_1228]
Hotel vicino San Giovanni


"La Via delle Api"
|
|
Esposizioni
|
Mostra: La Via delle Api. Prorogata fino a domenica 26 settembre 202125 Febbraio - 26 Settembre
- mostre -
|
Collezioni
Visite tattili-sensoriali
|
Video: “La Via delle Api”17 Giugno - 26 Settembre
- incontri -
|
Tassidermia
|
Video: l’Elefante “Toto” racconta la sua storia20 Luglio - 31 Dicembre
- incontri -
Video: il rarissimo esemplare di “Alca impenne” custodito al Museo Civico di Zoologia21 Luglio - 31 Dicembre
- incontri -
|






Tutti hotel a 5 stelle






























Tutti hotel a 4 stelle






























Museo
Missione
Il Museo Civico di Zoologia è un’Istituzione culturale che ha lo scopo di promuovere e diffondere le conoscenze scientifiche nell’ambito delle scienze biologiche e naturali.
Tradizionalmente, il cardine della sua attività è costituito dalla conservazione dei reperti zoologici che sottoposti a studi e indagini forniscono importanti informazioni per la ricostruzione della storia naturale e la conservazione dell’ambiente. Tutte le collezioni vengono accuratamente preservate ed incrementate. Ad oggi sono costituite da circa 4 milioni di esemplari e sono note in tutto il mondo scientifico per la loro importanza.
Tutti hotel a 3 stelle




























































L’attività di conservazione è affiancata da studi sulla sistematica, sulla biogeografia e sull’ecologia animale, testimoniati dalle numerose pubblicazioni scientifiche che annualmente vengono prodotte nei vari settori disciplinari.

Questo grande patrimonio di conoscenze, in continua crescita ed evoluzione, viene reso pubblico attraverso l’opera di divulgazione, educazione e formazione svolta sul territorio e presso la cittadinanza, per la quale il Museo ha istituito da circa vent’anni un Dipartimento educativo.
Famiglia hotel






























Animali domestici hotel






























Il primo obiettivo è stato costruire un rapporto con le scuole cittadine, che oggi frequentano in gran numero il Museo come parte integrante delle attività curricolari. L’attenzione è stata poi rivolta al pubblico comune. L’offerta del Museo si è arricchita di progetti culturali per tutti i tipi di visitatori (bambini, famiglie, anziani…), promuovendo la cultura scientifica e valorizzando nel contempo gli aspetti della fruizione sociale e dell’intrattenimento.
Hotels with pool






























Miglior boutique hotel hotel






























Negli anni è avvenuto anche un profondo cambiamento di impostazione culturale. È stata rivista l’immagine classica, autorevole e consolidata, del Museo “educatore” in favore di una struttura aperta a nuove esperienze culturali e al dialogo con la cittadinanza. In questo contesto si colloca l’interesse del Museo a rivestire un ruolo istituzionale moderno, adeguato alle necessità del territorio e alle sue problematiche socio-culturali.
Miglior hotel di lusso hotel






























Esposizioni

Hotel vicino Spagna





Hotel vicino Tiburtino





Amori bestialiLivree vistose, odori inebrianti e regali preziosi: l’arte della seduzione nel regno animale.
 |
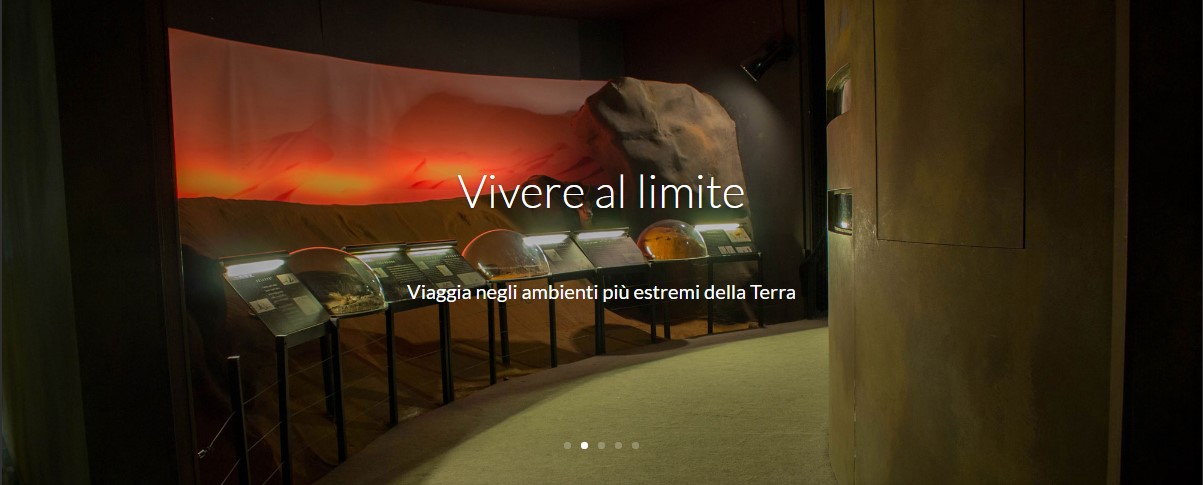
Vivere al limiteTemperature estreme, siccità e buio perenne: la sopravvivenza in condizioni limite.
 |
Barriera corallinaCoralli, tartarughe e pesci variopinti: un tuffo nella barriera corallina del Mar Rosso.
 |
Zone umide del LazioDa elefanti e leoni a lupi e cinghiali: un viaggio attraverso il tempo nella Campagna Romana.
 |
Sale Arrigoni degli OddiLa collezione di uccelli del Conte Ettore Arrigoni degli Oddi, padre dell’ornitologia italiana.
 |
Sale dei mammiferiI mammiferi e la loro diversità: alla scoperta di animali affascinanti, dalle forme a volte insolite e bizzarre.
 |
Sala di anfibi e rettiliSalamandre giganti, lucertole colorate e curiose tartarughe: il fascino “strisciante” di anfibi e rettili.
 |
Sala degli scheletriUna sala ammaliante e suggestiva dove mettere in gioco capacità di osservazione e riflessione.
 |
Sala della balenaMostre, convegni e molto altro ancora, per un incontro tra arte e scienza.
 |
Miglior hotel economico hotel






























Resort hotel






























Collezioni
Appartment



Tutti gli esemplari conservati hanno un grande valore scientifico perché rappresentano una testimonianza della biodiversità passata e presente. Sono continuamente sottoposti a studi, rilevazioni e indagini, aiutando gli scienziati a conoscere la storia naturale e fornendo importanti informazioni per la conservazione dell’ambiente.
Apart hotel

Gli zoologi curatori provvedono a conservare le collezioni in condizioni di sicurezza, ad incrementarle mediante la raccolta sul campo o attraverso donazioni e acquisti, ad inventariare i singoli esemplari, registrandone provenienza, numero e collocazione, e ne sono direttamente responsabili.
Bed and breakfast





I curatori sono affiancati dal prezioso lavoro del tassidermista del Museo, specializzato nella conservazione di animali a fini di studio ed esposizione.
GuestHouse





HolidayPark

ConchiglieCollezioni malacologiche
 |
InsettiCollezioni entomologiche
 |
InvertebratiCollezioni di invertebrati
 |
PesciCollezioni ittiologiche
 |
Anfibi e rettiliCollezioni erpetologiche
 |
UccelliCollezioni ornitologiche
 |
MammiferiCollezione teriologica
 |
Reperti osseiCollezione osteologica
 |
Hostel


Resort

Scienza
Tassidermia
Hotel vicino Tor Di Quinto



Hotel vicino Torrino

Hotel vicino Appio Latino


Le spoglie degli animali selvatici deceduti per incidenti o malattie vengono raccolte e portate al Museo da associazioni come il WWF e la LIPU, dai Parchi Naturali, dall’Istituto Zooprofilattico o anche da privati cittadini. La fase successiva è quella della preparazione per la conservazione mediante una tecnica chiamata tassidermia, dalle parole greche taxis=ordine e derma=pelle che significano mettere in posizione la pelle.
Hotel vicino Trastevere


È la sola pelle degli animali, infatti, con il pelo, le penne o le squame, ad essere sottratta alla naturale decomposizione mediante opportuni trattamenti.
Hotel vicino Ardeatino

Con questo procedimento di base si preparano le cosiddette “pelli da studio”, essenzialmente allo scopo di preservare nel tempo il materiale biologico e renderlo disponibile agli scienziati per le loro ricerche. Gli esemplari destinati all’esposizione subiscono una preparazione più complessa, detta naturalizzazione, che prevede il posizionamento della pelle su un modello artificiale ricostruito seguendo fedelmente le forme anatomiche dell’animale.
Hotel vicino Aurelio










Come risultato finale l’esemplare avrà un atteggiamento “naturale”, risvegliando nello spettatore curiosità ed interesse e rendendo evidenti alcuni concetti della zoologia in maniera molto più diretta di quanto riuscirebbero a fare parole scritte o immagini.
Hotel vicino Aventino

Hotel vicino Trevi

Nella tassidermia moderna, la conoscenza dell’anatomia e del comportamento animale, l’uso di materiali sempre più adeguati, la cura nei particolari e un notevole senso artistico concorrono alla realizzazione di preparati di grande valore scientifico ed estetico.
Hotel vicino Central Station



























Hotel vicino Trionfale







Hotel vicino Colosseum

Biblioteca
Hotel vicino Vaticano Prati



Il Museo Civico di Zoologia ha una biblioteca specializzata in biologia animale e conservazione della natura. Un settore della biblioteca è dedicato alle pubblicazioni moderne, mentre un settore storico raccoglie volumi e documenti antichi derivanti da donazioni e acquisizioni.
Hotel vicino Esquilino


Il settore moderno comprende circa 6500 monografie e varie riviste faunistiche; oltre a volumi strettamente zoologici, include importanti testi di geologia e paleontologia. Le monografie sono catalogate nel sistema informatico del Servizio Bibliotecario Nazionale (SBN), mentre le riviste non hanno ancora una catalogazione informatica.
Hotel vicino Via Veneto




Hotel vicino Villa Borghese Parioli





Rientra fra gli obiettivi del Museo rendere disponibile al più presto questo importante patrimonio di informazioni.
Hotel vicino Eur




Il settore storico ospita testi e documenti di grande valore come le monografie di Ulisse Aldrovandi, naturalista e museologo vissuto tra la fine del 1500 e l’inizio del 1600 e il fondo documentario del Conte Ettore Arrigoni degli Oddi, considerato da molti il padre dell’ornitologia italiana ed europea.
Hotel vicino Eur & Garbatella









Nel 1937, insieme alla collezione ornitologica del Conte, venne acquisita dal Museo anche la documentazione scientifica ad essa collegata, che include monografie e una miscellanea di importanti volumi e articoli, dei quali solo una parte attualmente catalogati (circa 1200 tra monografie ed articoli).
Hotel vicino Gianicolense






Del fondo fanno parte anche manoscritti e carteggi che raccolgono le pubblicazioni scientifiche di Arrigoni a partire dal 1883 (circa 1,5 m lineari).
Hotel vicino Giuliano-Dalmata
Allo stato attuale, la biblioteca del Museo Civico di Zoologia non dà in prestito i suoi volumi. È consentita la consultazione dei testi solo previo appuntamento.
Hotel vicino Magliana Vecchia
Le informazioni bibliografiche sul patrimonio librario conservato sono consultabili online presso il Polo SBN di Roma Capitale.
Hotel vicino Marsala


Hotel vicino Navona


Hotel vicino Nomentano







Visita |
Esplora |
Sito |
Hotel vicino Ostiense
